Effectively digest large or small dsDNA and ssDNA into mononucleotides
- Digest unwanted dsDNA and ssDNA molecules including small ssDNA such as random hexamers
- Use in sensitive applications requiring reverse transcription where any contaminating DNA is unwanted
Baseline-ZERO™ DNase* digests dsDNA and ssDNA into mononucleotides
more effectively than the commonly used bovine pancreatic DNase I. Even
the small DNA oligonucleotides that remain after treatment with bovine
pancreatic DNase I are undetectable by gel electrophoresis following
treatment with Baseline-ZERO DNase (Fig. 2). Removal of DNA from RNA
preparations is particularly beneficial when RNA in a sample is
amplified using a method that involves reverse transcription using
random primers, since any contaminating DNA would also be a template for
random-primed cDNA synthesis. 
Applications
- Removal of genomic DNA from small-sample total RNA preparations for expression analysis (Fig. 1)
- Removal of small DNA oligonucleotides (e.g., random primers)
| 
Figure 1. Real-time PCR of HeLa RNA preparations treated with various DNases. The lower the CT
value (intersection of curves with the red line), the greater the
amount of residual DNA not digested by the indicated DNase. Thus,
Baseline-ZERO™ DNase removed all detectable DNA from the RNA sample. The
TaqMan® probe assay amplified a 268-bp fragment of β-actin. Samples
were run in duplicate. |
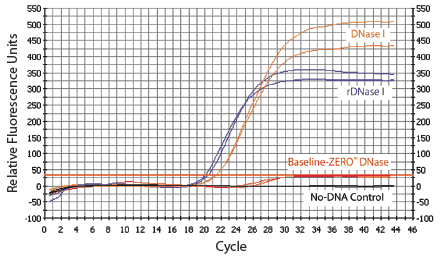

Figure 2: DNA treated with Baseline Zero is undetectable on standard gel electrophoresis.
160 ng of linear plasmid DNA was incubated for 15 minutes at 37°C as follows:
Lane M, Kilobase ladder
Lane 2, untreated;
Lane 3, DNase I treated;
Lane 4, Hyperactive DNase treated (supplier A);
Lane 5, Baseline-ZERO DNase treated.
Unit Definition: One Molecular Biology Unit (MBU) of Baseline-ZERO™ DNase produces an increase in the A260
of a solution of dsDNA, of 0.001 per minute at 25°C. Functionally, 1
MBU completely digests 1 µg of linear pUC19 DNA to mononucleotides in 10
minutes at 37°C.
Storage Buffer: Baseline-ZERO DNase is supplied in a 50% glycerol solution containing 50 mM Tris-HCl (pH 7.5), 10 mM CaCl2, 10 mM MgCl2 and 0.1% Triton® X-100.
10X Baseline-ZERO™ DNase Reaction Buffer: 100 mM Tris HCl (pH 7.5), 25 mM MgCl2 and 5 mM CaCl2.
10X Baseline-ZERO™ DNase Stop Solution: 30 mM EDTA.
Quality Control: Baseline-ZERO DNase is assayed for
its ability to completely digest linear dsDNA to mononucleotides under
standard assay conditions. Baseline-ZERO DNase is free of detectable
RNase activities as assayed by PAGE analysis of 1 µg of a synthetic RNA
transcript following an overnight incubation with sufficient DNase I to
completely digest 1,000 µg of DNA.
References
Kienzle, N. et al. (1996) BioTechniques 20, 612
If you cannot find the answer to your problem then please contact us or telephone +44 (0)1954 210 200